汕头大学医学院李康生、陈伟强和盛江涛联合团队提出转化生长因子β1可增强皮质神经元的放电活性
转化生长因子β1在调节细胞生长、分化、胚胎发生和损伤后修复中起着至关重要的作用[1]。一方面,转化生长因子β1是一种重要的神经保护因子,可促进神经元存活,并有助于减轻中枢神经系统基础疾病的病理过程,包括脑卒中、创伤性脑损伤、帕金森病和阿尔茨海默病[2-4]。另一方面,有报道称转化生长因子β1在新冠肺炎患者的阿尔茨海默病、轻度创伤性脑损伤和认知障碍中发挥神经毒性作用[3, 5, 6]。这些研究提示了转化生长因子β1在调节中枢神经系统生理和病理过程中的多效性。考虑到转化生长因子β1在中枢神经系统系统的多种病理过程中的作用,转化生长因子β1对神经元放电特性的影响及其分子基础仍需进一步研究。电压门控钠通道,负责产生动作电位的上升支,决定细胞膜的膜电势,并在神经递质的释放中发挥关键作用[7]。电压门控钠通道结构、功能或表达分布的改变会导致神经元兴奋性异常。多项研究报告称,转化生长因子β1可调节电压门控钠通道的功能和表达。在大鼠心房肌细胞中,转化生长因子β1会促进质膜上电压门控钠通道的缺失,降低电压门控钠通道电流,但在成年大鼠心室肌细胞中,转化生长因子β1会通过激活磷酸肌酸3-激酶(PI3K)/Akt通路促进电压门控钠通道电流的增加[8, 9]。在体外海马神经元创伤性损伤模型中,转化生长因子β1抑制电压门控钠通道电流和动作电位的产生[10]。这些研究表明,转化生长因子β1在调节可兴奋细胞电压门控钠通道方面具有多重作用,可能在心脏和特定脑区的可兴奋性细胞中发挥特定的作用。因此,研究病理条件下转化生长因子β1对皮质神经元电压门控钠通道的调控作用,有助于加深对中枢神经系统损伤机理的理解,并为后续相关的药物靶点开发提供建设性意见。
最近,来自中国汕头大学医学院李康生、陈伟强和盛江涛联合团队在《中国神经再生研究(英文版)》(Neural Regeneration Research)上发表了题为“Transforming growth factor-beta 1 enhances discharge activity of cortical neurons”的研究。该研究中利用转化生长因子β1处理小鼠原代皮质神经元,发现转化生长因子β1可能通过激活ERK,JNK和p38 MAPK信号通路,上调Nav1.3的表达,并以剂量和时间依赖性地增加皮质神经元电压门控钠通道的电流密度。此外,转化生长因子β1还能显著抑制小鼠皮质神经元的动作电位放电阈值。这一研究显示,转化生长因子β可通过激活MAPK通路导致Nav1.3表达增加,导致病理状态下皮质神经元动作电位的放电阈值降低,从而促进中枢神经系统神经兴奋性相关疾病的发生和进展,为未来的脑损伤治疗的策略起到一定的启发作用。
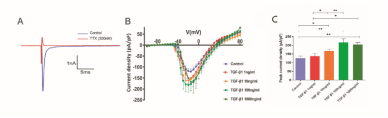
转化生长因子β1可通过激活PI3K-Akt通路,以剂量依赖的方式显著增加成年大鼠心室肌细胞中电压门控钠通道的电流密度[9],但却通过下调海马神经元的发射活动降低电压门控钠通道的电流密度[10]。这些结果证实了转化生长因子β1在不同兴奋细胞中调节电压门控钠通道功能的多效性。然而,转化生长因子β1对皮质神经元电压门控钠通道的作用仍然未知。为确定电压门控钠通道电流为河鲀毒素敏感电流,李康生、陈伟强和盛江涛联合团队用河鲀毒素(1 μM)处理神经元,随后检测电压门控钠通道电流。结果表明,河鲀毒素完全阻断了电压门控钠通道电流。结果提示,内向电流是由电压门控钠通道介导的河鲀毒素敏感电流。进而探讨转化生长因子β1是否引起电压门控钠通道电流增加,以及这种增加是否与转化生长因子β1浓度有关。用1-1000 ng/mL转化生长因子β1处理神经元24 h,从100 mV持电势开始逐级去极化诱导神经元内向电流,记录200 Hz下快速激活和失活。转化生长因子β1作用电压门控钠通道24 h后,电压门控钠通道的电流密度呈浓度依赖性增加,且1,10,100,1000 ng/mL时电压门控钠通道的电流密度分别为22.51%,30.50%,51.56%和41.89%。1 ng/mL转化生长因子β1刺激皮质神经元24 h对电压门控钠通道电流密度无明显影响,但随着转化生长因子β1浓度的增加,电压门控钠通道电流密度逐渐增加。在转化生长因子β1浓度为10-1000 ng/mL时,振幅较暴露前基线显著增加。-20 mV时的峰值电流也随转化生长因子β1浓度的变化而呈浓度依赖性(图1)。转化生长因子β1呈剂量依赖性地增加电压门控钠通道电流。
图1转化生长因子β1对培养小鼠皮质神经元电压门控钠电流密度的浓度依赖性影响
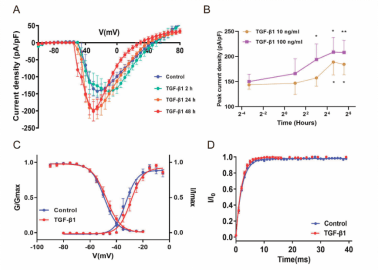
为探讨转化生长因子β1处理时间对皮质神经元电压门控钠通道电流密度的影响,李康生、陈伟强和盛江涛联合团队将原代皮质神经元暴露于转化生长因子β1(10 ng/mL)2-48 h,检测电压门控钠通道电流密度与转化生长因子β1暴露时间的关系。在转化生长因子β1作用下,Na+电流密度随时间增加,24和48 h时显著高于对照组。同样,在转化生长因子β1存在的情况下,-20 mV的标准峰值电流在24和48h也增加(图2)。结果显示,转化生长因子β1(10 ng/mL)作用于电压门控钠通道48 h后,逆转电压门控钠通道的能力呈下降趋势。为进一步了解转化生长因子β1对大脑皮质神经元电压门控钠通道动力学特征的影响。接下来研究了转化生长因子β1作用24h后神经元快速激活、稳态失活和恢复动力学特性(图2)。利用Boltzmann方程拟合了基于标准电流和预脉冲电压的电流-电压曲线。结果表明,转化生长因子β1不影响电压门控钠通道的稳态失活,电压门控钠通道恢复能力也没有明显影响,转化生长因子β1只改变了电压门控钠通道的快速活化特性。
图2转化生长因子β1诱导钠电流密度随时间的变化及Na+电流的电生理特性
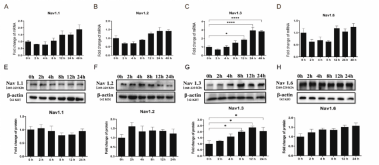
上述结果提示,转化生长因子β1可导致电压门控钠通道电流密度增加,而转化生长因子β1是否通过上调电压门控钠通道表达影响电压门控钠通道电流密度增加仍有待探究。因此,为探讨转化生长因子β1具体上调了那些亚型的电压门控钠通道,李康生、陈伟强和盛江涛联合团队对中枢神经系统中主要表达的钠通道亚型进行关注,通过PCR还有蛋白印迹方法,对Nav1.1,Nav1.2,Nav1.3和Nav1.6进行检测。结果显示,转化生长因子β1可显著性上调Nav1.3亚型的表达(图3)。结果提示,转化生长因子β1引起的Nav1.3亚型的表达上调可能是电压门控钠通道电流密度的增强的主导因素。
图3转化生长因子β1对皮质神经元电压门控钠通道表达的影响
在中枢神经系统损伤的情况下,Nav1.3是如何上调的,目前尚不清楚。细胞因子是一类调节神经炎症的分泌介质,可通过调节神经元的多种离子通道功能和神经元兴奋性参与创伤性脑损伤和脊髓继发性损伤的进展。如促炎细胞因子肿瘤坏死因子α通过激活JNK和p38 MAPK信号通路上调大鼠背根神经节神经元中的Nav1.3[11]。通过激活ERK信号通路也被证明能提高增强钠离子通道如Na1.7和Nav1.8的表达水平[12]。因此,李康生、陈伟强和盛江涛联合团队通过用PD98059(一种ERK1/2信号抑制剂),SP600125(一种JNK抑制剂)和SB203580(一种p38 MAPK抑制剂)对神经元进行预处理,以检测特定信号通路参与观察到的转化生长因子β1增强电压门控钠通道电流密度的能力(图4)。结果表明,转化生长因子β1可通过激活ERK,JNK和p38 MAPK信号通路上调皮质神经元中的Nav1.3,且不同的细胞因子可通过相同的信号通路促进神经元中的Nav1.3上调。
图4抑制MEK,JNK和p38 MAPK对转化生长因子β1诱导的皮质神经元钠电流密度和Nav1.3蛋白增强的影响
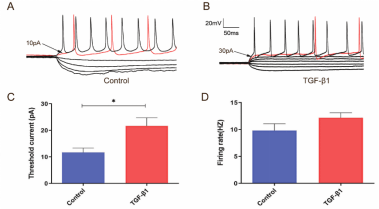
有报道称转化生长因子β1通过提高电压门控钾通电流密度来抑制海马和小脑颗粒神经元的放电活性,而电压门控钾通是形成动作电位下行分支的主要离子通道[10, 13, 14]。由此,实验验证了转化生长因子β1对大脑皮质神经元动作电位的影响。结果表明,转化生长因子β1可显著提高了神经元的放电阈值但却不改变其放电频率(图5)。结果进一步提示了转化生长因子β1可能对电压门控离子通道存在多重作用。然而,转化生长因子β1对皮质神经元电压门控钾通功能的影响仍需进一步验证。
图5 转化生长因子β1 对大脑皮质神经元动作电位的影响
综上所述,外源性转化生长因子β1以剂量和时间依赖的方式增强了培养小鼠皮质神经元的电压门控钠通道电流。这种增加与Nav1.3表达的激增有关,这可能是由涉及ERK,JNK和p38 MAPK信号通路的协同调节机制介导的。此外,转化生长因子β1可通过降低动作电位的放电值,显著增强小鼠皮质神经元的放电活性。这些结论提示,转化生长因子β1在病理状态下调节中枢神经系统神经毒性的方式。未来将在动物模型上进一步探究转化生长因子β1在创伤性脑损伤后不同阶段的作用,及其背后的具体机制,进一步明确转化生长因子β1在中枢神经系统损伤中的多重作用,为创伤性脑损伤个性化精准治疗提供理论依据。
原文链接:https://doi.org/10.4103/NRR.NRR-D-23-00756
参考文献
[1] Gomes FC, Sousa Vde O, Romão L. Emerging roles for TGF-beta1 in nervous system development. Int J Dev Neurosci. 2005;23(5):413-424.
[2] Li Z, Xiao J, Xu X, et al. M-CSF, IL-6, and TGF-β promote generation of a new subset of tissue repair macrophage for traumatic brain injury recovery. Sci Adv. 2021;7(11):eabb6260.
[3] Wyss-Coray T, Masliah E, Mallory M, et al. Amyloidogenic role of cytokine TGF-beta1 in transgenic mice and in Alzheimer's disease. Nature. 1997;389(6651):603-606.
[4] Oronsky B, Larson C, Hammond TC, et al. A review of persistent post-COVID syndrome (PPCS). Clin Rev Allergy Immunol. 2023;64(1):66-74.
[5] Patel RK, Prasad N, Kuwar R, et al. Transforming growth factor-beta 1 signaling regulates neuroinflammation and apoptosis in mild traumatic brain injury. Brain Behav Immun. 2017;64:244-258.
[6] Reiken S, Sittenfeld L, Dridi H, et al. Alzheimer's-like signaling in brains of COVID-19 patients. Alzheimers Dement. 2022;18(5):955-965.
[7] Simms BA, Zamponi GW. Neuronal voltage-gated calcium channels: structure, function, and dysfunction. Neuron. 2014;82(1):24-45.
[8] Ramos-Mondragón R, Vega AV, Avila G. Long-term modulation of Na+ and K+ channels by TGF-β1 in neonatal rat cardiac myocytes. Pflugers Arch. 2011;461(2):235-247.
[9] Kaur K, Zarzoso M, Ponce-Balbuena D, et al. TGF-β1, released by myofibroblasts, differentially regulates transcription and function of sodium and potassium channels in adult rat ventricular myocytes. PLoS One. 2013;8(2):e55391.
[10] Ondacova K, Moravcikova L, Jurkovicova D, et al. Fibrotic scar model and TGF-β1 differently modulate action potential firing and voltage-dependent ion currents in hippocampal neurons in primary culture. Eur J Neurosci. 2017;46(6):2161-2176.
[11] Chen W, Sheng J, Guo J, et al. Tumor necrosis factor-α enhances voltage-gated Na⁺ currents in primary culture of mouse cortical neurons. J Neuroinflammation. 2015;12:126.
[12] Yang F, Sun W, Yang Y, et al. SDF1-CXCR4 signaling contributes to persistent pain and hypersensitivity via regulating excitability of primary nociceptive neurons: involvement of ERK-dependent Nav1.8 up-regulation. J Neuroinflammation. 2015;12:219.
[13] Wulff H, Castle NA, Pardo LA. Voltage-gated potassium channels as therapeutic targets. Nat Rev Drug Discov. 2009;8(12):982-1001.
[14] Zhuang JL, Wang CY, Zhou MH, et al. TGF-β1 enhances Kv2.1 potassium channel protein expression and promotes maturation of cerebellar granule neurons. J Cell Physiol. 2012;227(1):297-307.
文章摘要:转化生长因子β1对中枢神经系统疾病的多效性已被广泛报道。在特定脑区,转化生长因子β1发挥的是神经保护还是神经毒性作用,可能取决于特定的病理过程和细胞类型。电压门控钠通道是神经元动作电位上升支中的一种重要的离子通道,与多种神经兴奋相关疾病有关。但是转化生长因子β1对电压门控钠通道功能特性和皮质神经元放电特性的影响尚不清楚。此次实验检测了转化生长因子β1对小鼠原代皮质神经元电压门控钠通道功能和放电特性的影响。结果发现,转化生长因子β1可通过上调Nav1.3表达,以剂量和时间依赖性的方式增强电压门控钠通道电流密度。然后通过用PD98059(ERK1/2抑制剂),SP600125(JNK抑制剂)和SB203580(p38 MAPK抑制剂)对神经元进行预处理,发现增加的电压门控钠通道电流密度和Nav1.3表达明显消除。有趣的是,转化生长因子β1还能显著增加皮质神经元动作电位的放电阈值,但未改变动作电位的放电速率。实验结果提示,转化生长因子β可通过激活ERK1/2-JNK-MAPK通路促进Nav1.3表达,导致病理状态下皮质神经元动作电位的放电阈值降低,从而促进中枢神经系统神经兴奋性相关疾病的发生和发展。
文章关键词:转化生长因子β1;皮质神经元;电压门控钠电流;Nav1.3;ERK;JNK;p38;放电活性;中枢神经系统;创伤性脑损伤
文章来源:Ren Z, Li T, Liu X, Zhang Z, Chen X, Chen W, Li K, Sheng J (2025) Transforming growth factor-beta 1 enhances discharge activity of cortical neurons. Neural Regen Res 20(2):548-556.
免责声明:如果侵犯了您的权益,请联系站长,我们会及时删除侵权内容,谢谢合作! |